
A week or so ago, Bruce and I derailed a simple question of winter collecting, and it would be more appropriate to take up the discussion in its own thread.
Some of our current research is focused around what ecologists call "resources partitioning". Theoretically, species using similar habitats must have "partitions" or differences in use among the resources available to avoid competing directly. We might simply call these "microhabitats" and the response that allows a particular species to dominate in that microhabitat can be morphological, behavioral, or both. The classic example is beak polymorphism in Darwin's finches:
http://en.wikipedia....arwin's_finches
Much of the work in community ecology has been done with birds, although there's a handful of studies that have investigated other vertebrate groups like small mammals, herps and reef fishes. In each case, limiting the routes of migration is critical to control, and so many of these studies have used "island-like" habitats such as oceanic islands, prairie dog colonies, desert seeps, coral reefs... Really unique habitats among the continuum of "environment".
If you know a little about darters, it may already be apparent why they are an ideal to investigate these processes... Many species live in riffles, which are somewhat of an island among the river continuum, and the dominant geologic characteristics are easily quantified. There are potentially multiple species and evolutionary lineages in the same riffle, there are a gob of different species to "assemble", and each has slightly different morphology. Males, females and juveniles have different patterning, so it's easy to discern - a member of one's own species may be the greatest competitor because they all need the exact same things. And individuals can only migrate in two directions... upstream and downstream. Furthermore, there's a bunch of unrelated species using the same habitats.
And most of all, we all know that darters just plain out rule
To conceptualize this, below is a graphic of a river in Ohio that has a pretty simple community. The darker colors for environmental variables are deeper and faster, each species has it's own color and number of individuals captured in one seine haul.
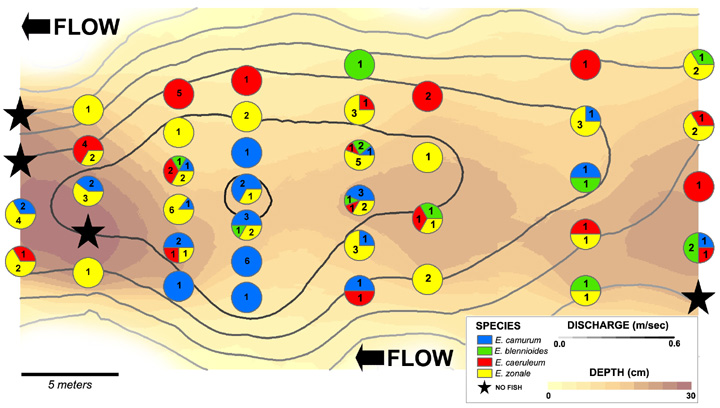
Figure 1
What you may notice is that while there is a ton of overlap, although each species occupies it's own space. This might be expressed in a conceptual model like this, where each ring is the species' distribution within this environment.
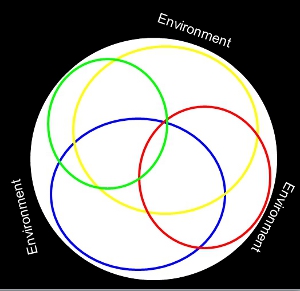
Figure 2
And then you take a bunch of samples at a bunch of different riffles and you can make something that looks like this to help visualize where species fall out in relation to one another:
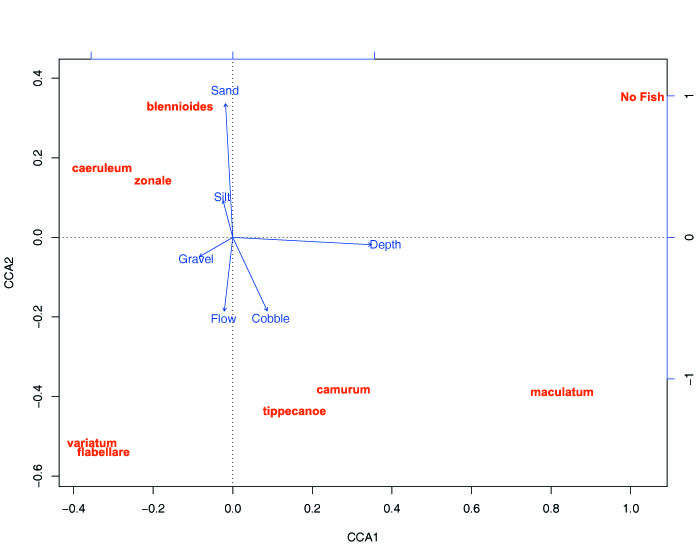
Figure 3
Each of the arrows are an environmental variable, the length of the arrow describes the strength of that variable in shaping the distribution of species and assemblages you are comparing. The distance of the species name from the center describes the average strength of association for that species to that variable. So, for example, zonale has a weak association to sand in silt compared to maculatum's strong association to depth and cobble. And samples taken in deep sandy places (the least riffle-like habitat) have no darters. You might also overlay the total distribution of any one species with "rings" like you see in Fig 2 to help see what's going on here.
Now... that's great. They live in different space at very local scales (such as a riffle or even a reach of river). There was a bunch of this work done in the 80's and 90's and people tried to come up with "rules of assembly", as though species distributions were some kind of fixed entity. Furthermore, a resulting assumption is that competitive have ended, it's all peace and love in darter communities, right?
But that's not what I observe, especially traveling the gradient from the Great Lakes into the species rich streams of Tennessee and over the hills to the darter deplete Atlantic Slope. I find that distributions are incredibly plastic (flexible) and are determined locally by both local geology and the potential pool of species.
To provide an anecdote to this effect, I had the fortunate chance of someone giving me an experiment. A year after I sampled the riffle in Fig 1, I returned to sample other riffles in the area. I felt the simplicity of this particular riffle was prime to take a high school teacher (with whom I was working) to better understand my work and have an enjoyable day in the field. A canoeist had placed a wing dam of cobble to cause a chute through the structure. This is the result:
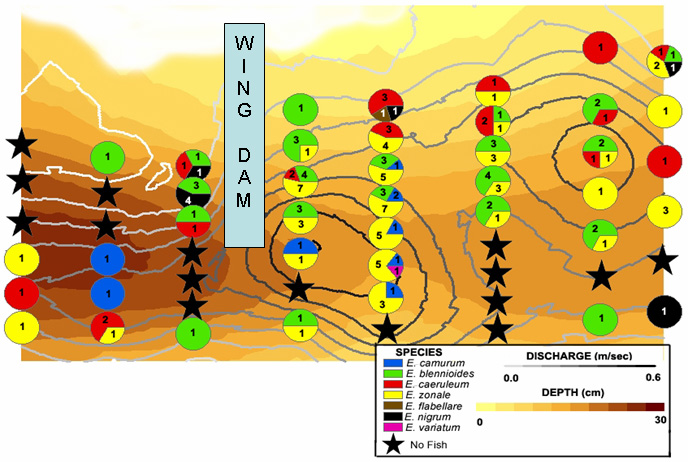
I am going to leave this to your interpretation for now... I think that's enough to digest in one post. Hopefully you'll find this engaging and we can get more deep into the discussion (I'm particularly eager to ask the question "What, then, is an invasive species?").
There are some things I will definitely address in consequent posts with Bruce in regards to his work and some pitfalls we've found... so regardless, this will continue whether you want it or not
Todd





